") 視覺系統(tǒng)的神奇之旅 視覺系統(tǒng)是怎么演化成今天樣子的?
視覺系統(tǒng)的神奇之旅 視覺系統(tǒng)是怎么演化成今天樣子的?
其實(shí),在動(dòng)物界,視覺系統(tǒng)亦經(jīng)歷了從簡(jiǎn)單到復(fù)雜的演化過程。對(duì)光敏感的結(jié)構(gòu)稱為光感受器(photoreceptor),它在單細(xì)胞動(dòng)物中可能只是探測(cè)周圍環(huán)境的明暗,而在多細(xì)胞動(dòng)物中則逐漸復(fù)雜化——出現(xiàn)了杯狀或囊狀光感受器,演化出晶狀體,可使光線聚焦,并形成清晰的圖像(image)。
人們認(rèn)為,這些圖像被轉(zhuǎn)化成一系列電信號(hào),再經(jīng)視神經(jīng)傳遞到視覺皮質(zhì)和其它腦區(qū)。據(jù)說(shuō),96%的動(dòng)物具有這種復(fù)雜的視覺系統(tǒng)(a complex optical system)(Land and Fernald 1992)。
一、視覺器官的演化
1.原生動(dòng)物的眼點(diǎn)
眼點(diǎn)(eyespot)是自然界中最簡(jiǎn)單的“眼睛”(Kreimer 2009),是存在于鞭毛蟲、能運(yùn)動(dòng)的綠藻細(xì)胞或其它像眼蟲這樣的單細(xì)胞光合生物中的一種光感受器官(photoreceptive organelle)。眼點(diǎn)由光感受器和亮的橙紅色色素顆粒區(qū)域所組成(圖1),這種光感受器是一種受藍(lán)光激活的腺苷酸環(huán)化酶(a blue-light-activated adenylyl cyclase)(Iseki et al. 2002),該受體蛋白的激發(fā)導(dǎo)致作為第二信使的環(huán)腺苷酸(cAMP)的形成,化學(xué)信號(hào)傳導(dǎo)最終觸發(fā)鞭毛拍打模式和細(xì)胞運(yùn)動(dòng)的改變。
簡(jiǎn)單地說(shuō),眼點(diǎn)使帶鞭毛的原生動(dòng)物能感受光的方向和密度,進(jìn)行定向的趨光運(yùn)動(dòng)或避光運(yùn)動(dòng),對(duì)這些小生命來(lái)說(shuō),這樣的功能足以滿足其生存需求了。據(jù)說(shuō),渦鞭毛蟲可通過眼點(diǎn)對(duì)光的感受來(lái)進(jìn)行捕食。
圖1扁眼蟲,紅色為眼點(diǎn)
2.刺胞動(dòng)物的感覺棒
水母是一種無(wú)脊椎動(dòng)物,歸屬于刺胞動(dòng)物門。水母眼睛中也有晶狀體和視網(wǎng)膜,其視網(wǎng)膜由色素構(gòu)成,但只是一種簡(jiǎn)單的板狀結(jié)構(gòu),當(dāng)然,這足以為它們提供光線強(qiáng)弱和方向的信息。令人驚訝的是,在視網(wǎng)膜上捕捉光線的感光細(xì)胞中,將光信號(hào)轉(zhuǎn)變成神經(jīng)電信號(hào)的蛋白質(zhì)以及向這種蛋白質(zhì)傳遞信號(hào)的環(huán)狀核苷是水母和人類都擁有的物質(zhì)。因此,可以推測(cè),人類的視覺系統(tǒng)有可能是從這種沒有大腦、相對(duì)原始的動(dòng)物祖先進(jìn)化而來(lái)。
生活在熱帶海洋中的一種稱為Tripedalia cystophora的箱水母,其傘帽頂部邊緣的兩側(cè)各有4個(gè)感覺棒(rhopalium),它們與4對(duì)觸手交替排列,晶狀體向內(nèi),指向傘帽的中央(圖2)。
每個(gè)感覺棒是四種不同形態(tài)眼的集群:一對(duì)凹眼(pit ocelli)、一對(duì)線眼(slit ocelli)、以及兩個(gè)不成對(duì)的透鏡眼(unpaired lens eyes),習(xí)慣上稱為大復(fù)眼(large complex eye)和小復(fù)眼(small complex eye),也可稱之為下透鏡眼(lower lens eye)和上透鏡眼(upper lens eye),上眼向上看,下眼水平看。這些眼由角膜、蜂窩狀透鏡和纖毛光感受體視網(wǎng)膜所構(gòu)成。
圖2一種箱水母(Tripedalia cystophora)及其視覺器官——感覺棒。A:水母?jìng)€(gè)體(比例尺=1cm);B:?jiǎn)蝹€(gè)感覺棒;C一個(gè)分離出來(lái)的感覺棒(比例尺=200μm)
這種水母有如脊椎動(dòng)物一樣復(fù)雜的光學(xué)系統(tǒng)(圖3),但還沒進(jìn)化出用于信息處理的中央大腦。而且它們的視網(wǎng)膜在焦點(diǎn)之外,表明清楚的圖像可能不是用來(lái)看清東西的,而是作為處理視覺信息的一種方式(Nilsson et al. 2005)。
水母眼睛的構(gòu)造雖然和脊椎動(dòng)物眼睛的構(gòu)造類似,但水母眼睛的晶狀體(lens)不能有效收集外界光線。隨著動(dòng)物的進(jìn)化,逐漸演化出杯狀或囊狀光感受器并形成可使光線聚焦的晶狀體。
圖3箱水母(Tripedalia cystophora)眼的精確解剖模型。矢狀面顯示有平衡石和兩個(gè)透鏡眼的內(nèi)部結(jié)構(gòu)。球形的晶狀體被蜂窩狀囊膜所包裹,其內(nèi)部在晶狀體和視網(wǎng)膜之間形成了類似于玻璃體的結(jié)構(gòu),大眼的虹膜收縮是由晶狀體囊的外面部分收縮引起的。下眼是回轉(zhuǎn)對(duì)稱的,但上眼僅兩側(cè)對(duì)稱(正面圖如右圖所示)。光感受器外節(jié)填滿了兩個(gè)透鏡眼的視網(wǎng)膜,光感受器外節(jié)的排列奇特,特別是在上眼中,在那里,受體軸(receptor axes)在透鏡一側(cè)的一個(gè)點(diǎn)上匯聚
3.環(huán)節(jié)動(dòng)物、軟體動(dòng)物和節(jié)肢動(dòng)物的復(fù)眼
這些無(wú)脊椎動(dòng)物的光感受器常常是一些紐扣狀的眼或凸出的視網(wǎng)膜,許多小眼排列在體表隆起之上,但仍位于小囊之內(nèi),小眼中的光感受細(xì)胞為色素所包圍,由于光線只能由一個(gè)方向進(jìn)入小眼,故而能感受光的方向。
昆蟲有稱之為復(fù)眼(圖4,圖5)的視覺器官,它由眾多的小眼(ommatidia)組成(如蝶、蛾類的復(fù)眼可含有約28000個(gè)小眼),每個(gè)小眼是一個(gè)獨(dú)立的感光單位,分別由角膜、晶椎、色素細(xì)胞、視網(wǎng)膜細(xì)胞、視桿細(xì)胞等所構(gòu)成。從視網(wǎng)膜細(xì)胞向后伸出的軸突穿過基膜匯合形成視神經(jīng)。通常每個(gè)小眼只能形成一個(gè)像點(diǎn),由眾多小眼的像點(diǎn)可拼成一幅圖像。
圖4昆蟲的復(fù)眼
圖5昆蟲復(fù)眼的解剖結(jié)構(gòu)示意圖
4.脊椎動(dòng)物的眼睛
脊椎動(dòng)物的視覺系統(tǒng)通常包括視網(wǎng)膜,相關(guān)的神經(jīng)通路和神經(jīng)中樞,以及為實(shí)現(xiàn)其功能所必須的各種附屬系統(tǒng)。這些附屬系統(tǒng)主要包括:眼外肌,可使眼球在各方向上運(yùn)動(dòng);眼(圖6)的屈光系統(tǒng)(角膜、晶體等),保證外界物體在視網(wǎng)膜上形成清晰的圖像。
圖6人的眼睛(來(lái)源:維基百科)
二、光感受器的演化
所有的光感受器都具有對(duì)光敏感的色素(如類胡蘿卜素、視紫紅質(zhì)),這些色素吸收光能的光子,再產(chǎn)生發(fā)生器電位(generator potential)。
一些含有葉綠素的鞭毛類原生動(dòng)物(單細(xì)胞生物)具有大量的亮紅色光感受器顆粒,稱之為眼點(diǎn)(stigma)(圖7a)。這種顆粒是類胡蘿卜素,但實(shí)際上的光感受器就是鞭毛基部的膨大物。眼點(diǎn)可能起到了一種保護(hù)作用,如果光感受器要探測(cè)從某個(gè)方向而不是別的方向來(lái)的光的話,眼點(diǎn)就是必須的。因此,光感受器加上眼點(diǎn)可使眼蟲定位自身以便光感受器可以感受到光。這可使原生動(dòng)物定位于具有足夠的水柱中光以滿足自身光合作用的需求(Miller and Harley 2001)。
一些無(wú)脊椎動(dòng)物,如蚯蚓類的正蚓(Lumbricus),具有簡(jiǎn)單的單細(xì)胞的光感受器細(xì)胞,分散在表皮上或集中在身體的特定區(qū)域。另有一些無(wú)脊椎動(dòng)物,如扁蟲,具有稱之為單眼(ocelli)的結(jié)構(gòu),呈小杯狀,排列有一些光敏感受器以及一些吸光色素(圖7b)。這種光敏細(xì)胞叫做小網(wǎng)膜細(xì)胞(retinular cell),含有光敏色素。
光的刺激引起色素的化學(xué)變化,導(dǎo)致產(chǎn)生發(fā)生器電位,這將激發(fā)感覺神經(jīng)元攜帶的可在身體的其它地方解釋的動(dòng)作電位。這種類型的視覺系統(tǒng)給動(dòng)物提供光的方向和密度,但不形成圖像。單眼普遍存在于環(huán)節(jié)動(dòng)物、軟體動(dòng)物和節(jié)肢動(dòng)物等門類。
復(fù)眼由數(shù)個(gè)或許多小眼組成。雖然復(fù)眼也見于一些環(huán)節(jié)動(dòng)物和雙殼類軟體動(dòng)物,但在節(jié)肢動(dòng)物中最為發(fā)達(dá),也被了解得最為清楚。由于每個(gè)小眼在方位上略有不同,就形成了眼睛在整體上凸出的形狀(圖7c)。復(fù)眼的視野非常寬,就像我們每個(gè)人曾經(jīng)想抓住蝴蝶時(shí)都會(huì)有這樣的感覺。
每個(gè)單眼都有自己的神經(jīng)束,匯集成了一個(gè)大的視神經(jīng),相鄰單眼的視野也有一些相互重疊。此外,這些無(wú)脊椎動(dòng)物也具有相當(dāng)復(fù)雜的中樞神經(jīng)系統(tǒng)。因此,復(fù)眼在探測(cè)運(yùn)動(dòng)方面非常有效,也可能能形成圖像。此外,許多復(fù)眼也能適應(yīng)光密度的變化,一些具有彩色視力。
烏賊和章魚的復(fù)雜相機(jī)狀眼是無(wú)脊椎動(dòng)物中最好的形成圖像的眼睛。烏賊的眼巨大,是動(dòng)物中最大的眼,直徑超過了38cm。頭足類動(dòng)物的眼睛常可與脊椎動(dòng)物的眼睛相媲美(圖7d),因?yàn)樗鼈兙哂幸粋€(gè)薄的、透明的角膜和一個(gè)晶狀體可聚焦視網(wǎng)膜上的光,而且被睫狀肌所懸掛與控制。
在脊椎動(dòng)物的眼中,視網(wǎng)膜層是倒置的,感受體是視網(wǎng)膜的最深的細(xì)胞,兩只眼都聚焦并形成圖像,雖然過程的細(xì)節(jié)不同。在陸生脊椎動(dòng)物中,改變晶狀體形狀(厚度)的肌肉聚焦光線,而在魚類和頭足類中,光線聚焦的方式是通過肌肉使晶狀體面向或離開視網(wǎng)膜進(jìn)行運(yùn)動(dòng)以及改變眼球的形狀來(lái)實(shí)現(xiàn)的。
圖7無(wú)脊椎動(dòng)物的光感受器:(a)原生動(dòng)物眼蟲的眼點(diǎn),(b)扁蟲的單眼,(c)昆蟲的復(fù)眼,(d)復(fù)雜的相機(jī)狀眼(引自:Miller and Harley 2001)
三、視覺分子的起源
對(duì)生命來(lái)說(shuō),沒有什么比光更重要的了,它在能量上的重要性無(wú)可比擬。對(duì)動(dòng)物來(lái)說(shuō),雖然能量重要性是間接的,但光卻是它們行為的探照燈,并且借用了古菌中的一種重要的光能感應(yīng)分子——視黃醛(retinene)作為它們視覺的開關(guān)。
1.眼睛中的神奇物質(zhì)——視黃醛
在我們眼睛的視網(wǎng)膜中,有一類與感光有關(guān)的色素蛋白復(fù)合體(稱之為視紫紅質(zhì),rhodopsin),其核心的分子就是視黃醛,它通過光的介導(dǎo)及自身構(gòu)象的變化推動(dòng)了視紫紅質(zhì)的分解與合成,形成了所謂的視覺循環(huán)(visual cycle)。一種異構(gòu)體——11-順視黃醛與視蛋白的一個(gè)賴氨酸以共價(jià)鍵相結(jié)合,在黑暗中穩(wěn)定,但在光照條件下,轉(zhuǎn)變成另一種異構(gòu)體——全-反視黃醛(圖8),從視蛋白脫落,造成視紫紅質(zhì)的漂白,這也稱之為視網(wǎng)膜的光分解反應(yīng)。
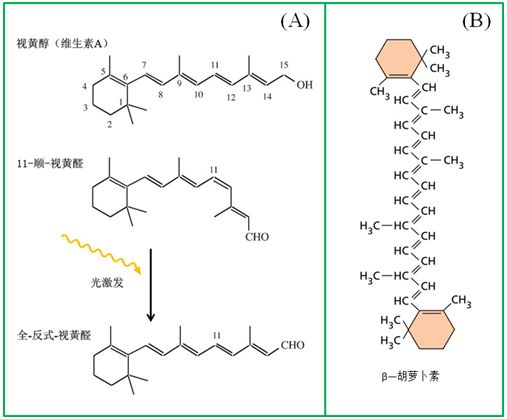
圖8視覺的分子起源。(A)在光的驅(qū)動(dòng)下,視黃醛在反式與順式異構(gòu)體之間的切換;(B)與視黃醇(醛)之間在結(jié)構(gòu)上具有同源性的β—胡蘿卜素
視紫紅質(zhì)是一種特殊的光感受體,強(qiáng)烈地吸收藍(lán)綠光而呈現(xiàn)紅紫色,這就是為何叫做“visual purple(視紫紅)”的緣故,它負(fù)責(zé)在黑暗中的單色視覺(Stuart and Brige 1996)。每個(gè)視桿細(xì)胞中大約含有1千萬(wàn)個(gè)視紫紅質(zhì)分子,分布在細(xì)胞外段由細(xì)胞膜折疊而成的1千個(gè)膜盤上(圖9)。
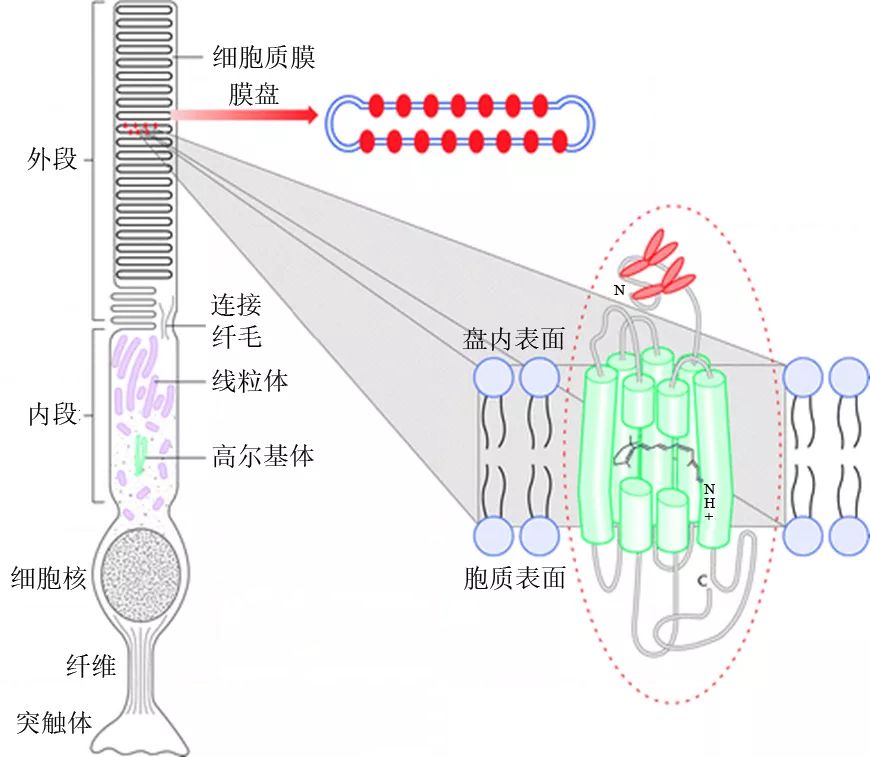
圖9視桿細(xì)胞與分布于其膜盤上的視紫紅質(zhì)分子(來(lái)源:http://scientifact.blogspot.com/2014/08/how-do-we-sense-light-rhodopsin.html)
視紫紅質(zhì)分為I型和II型,I型視紫紅質(zhì)最早發(fā)現(xiàn)于一類古老的原核生物——極端嗜鹽古菌Halobacterium salinarum中,在這種細(xì)菌中含有四種視紫紅質(zhì)—Bacteiorrhodosin(Br,質(zhì)子泵)、Halorhodopsin(Hr,氯離子泵)及兩種Sensoryrhodopsin(SR I和II),II型視紫紅質(zhì)主要分布于高等真核生物,包括動(dòng)物眼睛中的視紫紅質(zhì)、人類的桿細(xì)胞和錐細(xì)胞的視覺色素、低等脊椎動(dòng)物的松果體和丘腦等中(王寧等2010)。
2.在嗜鹽古菌中視黃醛可用于生產(chǎn)ATP
嗜鹽古菌卻可以利用視黃醛來(lái)生產(chǎn)ATP。這種微生物通過光誘導(dǎo)視黃醛的結(jié)構(gòu)變化來(lái)推動(dòng)質(zhì)子的跨膜傳送,即視黃醛通常以順式存在于紫膜(細(xì)胞膜上的斑狀紫色膜片組織)內(nèi)側(cè),它可被光激發(fā)轉(zhuǎn)換成全—反式結(jié)構(gòu)狀態(tài),其結(jié)果使H+轉(zhuǎn)移到膜外,隨著視黃醛分子的松弛和質(zhì)子的吸收,又轉(zhuǎn)變回較穩(wěn)定的順式狀態(tài),又再次被光激發(fā),轉(zhuǎn)移H+,如此循環(huán)(圖8A)。正是這種跨膜的H+梯度可在ATP酶的催化下完成ATP的合成。
對(duì)光養(yǎng)或無(wú)機(jī)化養(yǎng)微生物來(lái)說(shuō),跨膜質(zhì)子梯度和ATP合成酶是ATP生成的必要條件。放氧光合生物通過水的裂解(類囊體膜內(nèi))既提供了電子,也釋放出H+從而形成了跨膜H+梯度,使ATP合成酶利用跨膜的質(zhì)子動(dòng)力勢(shì)將ADP轉(zhuǎn)化成ATP,在這一過程中釋放到類囊體膜外的H+又可與高能的電子一道用于形成還原力—NADPH,這也解決了H+在細(xì)胞中的堆積問題。顯然,光合放氧生物(藍(lán)細(xì)菌)的ATP生產(chǎn)模式(與復(fù)雜的光系統(tǒng)I和II偶聯(lián))得到了自然界青睞,而與視紫質(zhì)偶聯(lián)的相對(duì)簡(jiǎn)單的ATP的生成方式并不普遍。
3.同樣的感光物質(zhì)服務(wù)于完全不同的生存目的
視紫紅質(zhì)廣泛存在于各種古菌和真細(xì)菌之中,但令人驚訝的是,它也廣泛存在于動(dòng)物的視覺細(xì)胞中。為何動(dòng)物視覺借用了這種古菌和細(xì)菌中的感光物質(zhì)(視黃醛)而不是其它的感光物質(zhì)(如葉綠素、類胡蘿卜素等)?此外,視黃醛從何而來(lái)呢?只要比較一下圖8A與圖8B就不難看出β—胡蘿卜素和視黃醇(醛)之間在結(jié)構(gòu)上的同源性,事實(shí)上,β—胡蘿卜素在氧化酶催化下可裂解為2分子視黃醇(醛),視黃醇在醇脫氫酶作用下轉(zhuǎn)化為視黃醛。
顯然,在結(jié)構(gòu)上同源的感光物質(zhì)在進(jìn)化的過程中被不同的生物類群用于了完全不同的生存目的:在放氧光合生物中,類胡蘿卜素參與光能的吸收和傳遞,并可淬滅激發(fā)態(tài)的葉綠素而保護(hù)后者;在一些古菌中,其氧化產(chǎn)物—視黃醛與視蛋白結(jié)合成視紫質(zhì),利用光能形成跨膜的H+梯度為ATP的合成創(chuàng)造條件,推動(dòng)光能向化學(xué)能的轉(zhuǎn)變;而在動(dòng)物的視覺細(xì)胞中,視紫質(zhì)通過復(fù)雜的生化反應(yīng)可將光能轉(zhuǎn)化成為神經(jīng)信號(hào)。無(wú)論這是一種偶然還是偶然中的必然,都與光有關(guān),只是從能量轉(zhuǎn)換走向了信息(視覺)感知。
-
發(fā)生器
+關(guān)注
關(guān)注
4文章
1368瀏覽量
61677 -
感受器
+關(guān)注
關(guān)注
0文章
12瀏覽量
3409
原文標(biāo)題:視覺系統(tǒng)的演化之旅——視覺器官、光感受器及視覺分子
文章出處:【微信號(hào):AItists,微信公眾號(hào):人工智能學(xué)家】歡迎添加關(guān)注!文章轉(zhuǎn)載請(qǐng)注明出處。
發(fā)布評(píng)論請(qǐng)先 登錄
相關(guān)推薦
安森美機(jī)器視覺系統(tǒng)解決方案
Zebra GevIQ榮獲2024《視覺系統(tǒng)設(shè)計(jì)》創(chuàng)新獎(jiǎng)
機(jī)器視覺系統(tǒng)用途與實(shí)現(xiàn)及市場(chǎng)趨勢(shì)
機(jī)器視覺系統(tǒng)如何選擇圖像傳感器
CCD機(jī)器視覺系統(tǒng)的工作原理和特性
機(jī)器視覺系統(tǒng)與運(yùn)動(dòng)控制系統(tǒng)的區(qū)別
機(jī)器視覺系統(tǒng)的工作原理和應(yīng)用領(lǐng)域

機(jī)器視覺系統(tǒng)五個(gè)模塊介紹

機(jī)器視覺系統(tǒng)的組成部分

海伯森攜其最新技術(shù)成果亮相武漢VisionCon視覺系統(tǒng)設(shè)計(jì)技術(shù)會(huì)議
視覺系統(tǒng)所使用的相機(jī)種類介紹

機(jī)器人視覺系統(tǒng)深度解析
機(jī)器視覺系統(tǒng)和人工智能有什么區(qū)別
機(jī)器視覺系統(tǒng)中常用攝像機(jī)的分類






 工商網(wǎng)監(jiān)
工商網(wǎng)監(jiān)
評(píng)論